Planta herbácea de clima quente e úmido, originária da Índia, a maconha (Cannabis sativa) pertence à família Moraceae. Farmacologicamente, o principal constituinte psicoativo desse tipo de planta é o tetrahidrocanabinol (THC), um dos 400 compostos da planta, incluindo outros canabinoides, como o canabidiol (CBD), canabinol(CBN), e tetrahidrocanabivarin (THCV). O THC é semelhante amandamida, que é um composto presente pelo organismo de alguns animais.
Quando ocorre a inalação da fumaça, a Cannabis pode produzir vários efeitos subjetivos em humanos: euforia, disforia, sedação, alteração da percepção do tempo, aumento da interferência na atenção seletiva e no tempo de reação, alteração nas funções sensoriais, prejuízo de controle motor, do aprendizado e prejuízo transitório na memória de curto prazo, além de efeitos neurovegetativos como boca seca, taquicardia e hipotensão postural. Efeitos adversos incluem crises de ansiedade, ataques de pânico e exacerbação de sintomas psicóticos existentes.
Os produtos naturais encontrados na maconha ligam-se aos receptores de canabinóides, que são receptores acoplados a proteína G cujo ligante endógeno é a amandamida, um derivado do ácido araquidônico. Os dois receptores de canabinóide conhecidos são denominados.CB1 e CB2. A anandamida possui propriedades bioquímicas e farmacológicas semelhantes ao THC, encontrado na maconha.
Estrutura química da Anandamida e do 2-glicerol araquidonii (2AG)
Fonte: Honório et all, 2004
Os receptores canabinóides CB1 estão distribuídos em muitas áreas do sistema nervoso central, principalmente nos gânglios basais, cerebelo, hipocampo, córtex, amígdala e núcleo acumbens. Já os receptores CB2 estão localizados em áreas periféricas como, por exemplo, células do sistema imunológico. Em relação aos demais neurotransmissores, seus níveis no cérebro são semelhantes aos de dopamina e serotonina, mas bem menores que os de GABA e glutamato.
Assim, ao inalar a fumaça da maconha, o THC vai diretamente para alvéolos sendo absorvidos facilmente. Minutos depois de inalado, o THC cai na corrente sanguínea, chegando até o cérebro. O THC se liga aos receptores canabinoides e ativa os neurônios, influenciando de forma adversa o cérebro. A interação do THC com o cérebro pode causar sentimentos relaxantes, como sensação de leveza, sendo que outros sentidos também podem se alterar.
A Ampulex compressa (vespa-joia)é um
parasita específico da barata Periplaneta americana, o ataque e a injeção do veneno da vespa na barata se dar por movimentos precisos e coordenados, o que garante a completa inanição da barata, garantindo o desenvolvimento das larvas de Ampulex dentro do corpo desta.
O ataque é composto por duas picadas. A primeira dá-se no Gânglio torácico, cujo veneno é injetado pela primeira vez, ocasionando à perda do movimento das pernas dianteiras da barata. O veneno da vespa é composto por grandes quantidades de acido gama-amino-bultríco (GABA), Taurina e Beta-alanina. A mistura bloqueia temporariamente os potencias de ação dos movimentos da barata, através do declínio da ação dos canais de sódio e cloro, comprometendo a condução iônica através do axônio, resultando na perda das sinapses nervosas. Individualmente, estas substâncias induzem o estado de paralisia da barata. A ação dessas em conjunto se mostra mais eficazes, o GABA é responsável pela ativação dos canais de Sódios e Cloro por ligar-se aos receptores GABA, nos neurônios, a Taurina e a Beta-alanina prolongam a duração do efeito paralisante, diminuindo a absorção do GABA na fenda sináptica. Combinados, esse coquetel de substancias paralisa e impede a barata de mover-se e defender-se enquanto a vespa se prepara para a segunda picada.
A segunda picada se dá em locais precisos no gânglio subesofágico (SEG) situado próximo à cabeça da barata, na seção que controla a fuga e o reflexo. As neurotoxinas do veneno da vespa causam um bloqueio na transmissão sensitiva da substância responsável pelo movimento, a dopamina. A dopamina é um neurotransmissor sintetizado em áreas do sistema nervoso central e periférico, e assim que formada, é encapsulada em vesículas para posteriormente ser liberada durante a sinapse, ela possui a função ligada ao movimento, a memória, a atenção, a recompensa agradável, dentre outras. Tais vesículas encontram-se concentradas no terminal axônico, e quando os impulsos nervosos chegam a esse terminal, os neurotransmissores são liberados por meio da exocitose. Para ocorrer a sinapse é necessária a transferência dos neurotransmissores da membrana pré-sináptica para a membrana pós-sináptica. Como resposta a tal, a barata primeiro irá se debater, e em seguida, esses movimentos tornarão-se mais lentos até a mesma não apresentar respostas normais de defesa e fuga. A ação dessa segunda picada se dá, também, sobre o neurotransmissor octopamina, bloqueando seus receptores – estes responsáveis pelo controle dos movimentos e sensações (semelhante a dopamina, em humanos). Ao bloquear a octopamina o veneno da vespa jóia, impede que a sinapse química ocorra ao impossibilitar que os impulsos nervosos passem de um neurônio pré-sinático para o neurônio pós-sinático na fenda sináptica por meio do neurotransmissor químico, semelhante a dopamina. Desse modo, os componentes do veneno afetam diretamente os neurônios no cérebro que modulam as sinapses no tórax, o que faz com que a barata primeiramente apresente um comportamento de limpeza, e em seguida entre em um estado que é caracterizado por pouca, ou nenhuma, atividade locomotora espontânea o que torna o metabolismo lento e insensível a diversos estímulos.
Transferência dos neurotransmissores da membrana pré-sináptica para a membrana pós-sináptica.
Deste modo a barata fica a mercê da vespa, que a carrega para a sua toca, deposita seu ovo na axila da barata, e dali eclodirá em poucos dias uma larva que se alimentará da barata viva, porem em estado de metabolismo basal (zumbi), imóvel e indefesa, até que todos os seus órgãos sejam consumidos pela larva e esta desenvolva-se em uma nova vespa joia.
Gal, Ram, Lior Ann Rosenberg, and Frederic Libersat. “Parasitoid wasp uses a venom cocktail injected into the brain to manipulate the behavior and metabolism of its cockroach prey.” Archives of Insect Biochemistry and Physiology 60, no. 4 (2005): 198-208. doi:10.1002/arch.20092.
Remédio ou veneno? Uma substância extraída de um sapo da Amazônia vem sendo usada como um suposto remédio pra várias doenças, porém não se tem estudos ainda que comprovem se todas as substâncias existentes neste veneno possuem eficácias na medicina.
A Phyllomedusa bicolor, conhecido popularmente como "Sapo Kambô", é um anfíbio da família Hylidae, famílias das pererecas, encontrado na Amazônia. É conhecido devido a utilização do seu veneno na medicina tradicional. É a maior espécie do gênero, podendo chegar a 11,8 cm de comprimento e também um dos maiores hilídeos da Amazônia.
O veneno do Kambô é utilizado pelos indígenas como uma forma de medicina natural e acreditam que este veneno fortalece o sistema imunológico e afastar o "Panema" - má sorte. Com a colonização no norte, os seringueiros aprenderam a usar essa técnica e atualmente a aplicação da vacina-do-sapo já ocorre em várias regiões do Brasil. Para aplicar a vacina-do-sapo é necessário queimar o braço e tórax (homens) ou perna (mulheres) com uma ponta de um pequeno cipó fazendo vários pontos. O veneno é então aplicado nessas pequenas queimaduras. Os sintomas são quase imediatos e a pessoa sente um forte calor, náuseas, dor no estômago, vômitos e outros desconfortos. A ação do veneno dura em média 15 minutos.
A pele dos anfíbios exerce mais funções além de ser uma estrutura de revestimento, desempenhando principalmente o transporte de água e solutos, auxiliando na respiração, regulação da temperatura corporal e defesa contra o ataque de microorganismos e predadores. Apresentam duas glândulas, uma mucosa que secreta muco para manter a umidade da superfície do muco favorecendo a respiração cutânea; outra granulosa, frequentemente é tóxica ou repelente, é responsável pela defesa passiva dos anfíbios. A secreção das glândulas granulosas apresenta um aspecto viscoso e leitoso e ocorre, normalmente após estresse ou injúria. Esta glândula é controlada por inervação simpática e seu conteúdo é secretado sob regulação neuronal, possivelmente neuropeptídeos.
As glândulas granulosas ou de veneno são responsáveis pela síntese e armazenamento de uma grande diversidade de compostos químicos que apresentam propriedades antibióticas e atuam na defesa de microorganismos. Quimicamente, esses compostos, podem ser agrupados em cinco classes: aminas biogênicas, peptídeos, proteínas proteolíticas, alcaloides e esteróides. Estudos feitos com as secreções obtidas da pele de anfíbios, indicaram que representantes das famílias de Hylidade apresentam compostos de aminas, peptídeos e proteínas.
Ocorrência de compostos bioativos encontrados entre os representantes da anfibiofauna: (+) presente, (-) ausente, (o) ocorrência não avaliada.
As aminas biogênicas são mediadores químicos bastante comuns entre os animais. Apresentam estrutura química e ações farmacológicas bastante diversificadas. Uma dessa estrutura química encontrada na família desse anfíbio é a amina indólica que tem a atividade farmacológica ligada a ação estimulante em musculatura lisa. Assim, os músculos lidos extravasculares são contraídos pela ação dessas substâncias. Podem alterar, também, a função motora do trato gastrointestinal, termorregulação e induz a secreção de catecolaminas pelas glândulas supra-renais. As catecolaminas são representadas pela epinefrina, norepinefrina, dopamina e epinina e apresentam uma ação vasopressora potente causando um aumento nas taxas cardíacas e respiratória. Em altas taxas podem causar espasmos, vômitos e convulsões.
Outra amina encontrada na secreção dos representantes de Hylidae foi às histaminas que podem provocar reações alérgicas como urticária, irritação nas mucosas nasais e oculares, lacrimejamento e espirros.
Estrutura química das aminas encontradas nos anfíbios.
As substâncias mais abundantes encontrada nos indivíduos de Phyllomedusa bicolor são os peptídeos com características vasoativas -adenoregulina, além desta também foram encontrada substâncias como os peptídeos opióides. Estas substâncias podem provocar um estado de letargia que supera os efeitos da morfina ou apatia duradoura em ratos. Nos humanos não há ainda estudos que comprovem os efeitos causados pelos peptídeos opióides.
Estes peptídeos apresentam cargas positivas, podendo formar hélices anfifílicas de comprimento variando de um peptídeo para outro, possuindo habilidade de se autopolimerizar formando longos filamentos com os grupamentos hidrofílicos para o interior e os hidrofóbicos em contanto com as partes lipofílicas da membrana.
Em resumo a parte positiva do peptídeo interage com os fragmentos lipídicos, induzindo a formação de alfa-hélice e polimerização. O complexo supramolecular em alfa-hélice liga-se superficialmente a região hidrofóbica e começa a penetrar a membrana permitindo a entrada dos íons de cloro e o efluxo de hidroxilas e potássio despolarizando a membrana e provocando um potencial de ação. Assim, os indivíduos de Phyllomedusa bicolor obtêm sucesso para seu propósito - a proteção contra indivíduos. Agindo desta forma o veneno desacopla a respiração dos microorganismos resultado na morte destes.
Ataques de tubarões podem parecer cruéis mas, é importante salientar que eles não são criaturas assassinas. Os tubarões, como qualquer outro animal, seguem seus instintos para se alimentarem e se defenderem. A dieta de um tubarão consiste principalmente peixes, tartarugas marinhas, baleias, leões-marinhos e focas e outros animais marinhos com grande teor de gordura.
Os tubarões pertencem a classe dos Condrictes e à superordem dos Selachimorpha e são assim clasificados por apresentarem um esqueleto cartilaginoso, que faz com que o corpo seja mais é flexível tendo cerca de metade da densidade do osso, reduzindo o peso do esqueleto e poupando energia, e um corpo hidrodinâmico.
Para a efetividade dos seus ataques os tubarões apresentam dentes não incorporados no maxilar e que são constantemente trocados ao longo da vida do animal e são distribuídos em diversas fileiras que crescem na parte interna da mandíbula e maxila. Para aumentar a eficácia da mordida, os dentes são serrilhados.
Além dos dentes especializados para caça, os tubarões podem sentir o cheiro de sangue a quilômetros de distância, e, provavelmente, ocorre devido a ao formato e fisiologia do nariz. Assim, conforme o tubarão nada, a água flui através de duas narinas frontais. A água entra pela passagem nasal e passa por dobras de pele cobertas por células sensoriais. As narinas são dois orifícios localizados na região cefálica, que terminam em fundos cegos. Não se comunicam com a faringe, como ocorre na maioria dos peixes ósseos, assim, as narinas têm apenas a função olfativa: um dos sentidos muito desenvolvidos nos tubarões. O odor é percebido por quimiorrecepção: um dos mecanismos sensoriais utilizados para a percepção de presas a grandes distâncias
As narinas são ricas em terminações nervosas do nervo olfativo. Os dendritos das células olfativas possuem prolongamentos sensíveis. Assim, quando alguma partícula olfativa entra pelas narinas do tubarão, entra em contato com esses prolongamentos sensíveis geram impulsos nervosos, que são conduzidos até o corpo celular das células olfativas, onde atingem os axônios, que se comunicam com o bulbo olfativo. O estímulo é estão levado ao Sistema Nervoso Central (SNC), onde o processo de sinalização é interpretado e decodificado.
Além do olfato aguçado, os tubarões tem uma visão consideravelmente boa. Alguns pesquisadores acreditam que a lente dos tubarões está suspensa por um ligamento dorsal, estando adaptada à visão a distância; para a uma visão mais próxima ela é movida para frente pela tração de um pequeno músculo protrator, fixo à lente. Seus olhos por ficarem nas laterais da cabeça podem ampliar seu campo de visão para quase 360°. Entretanto, por estarem separados, os tubarões não tem noção de profundidade, possuindo uma visão mais plana.
Os olhos possuem uma camada refletiva, a qual permite um aproveitamento superior da luminosidade em locais com pouca luz, como as águas turvas ou profundas e à noite. A essa estrutura da-se o nome de tapetum lucidum. O tapetum lucidum é uma agrupamento de células que contêm cristais de guanina e estes cristais agem como espelhos refletindo a luz de volta para a retina, aumentando as chances de uma maior absorção luminosa.
Apresentam também uma grande sensibilidade às vibrações devido ao ouvido interno, responsável pelo equilíbrio e detecção das vibrações de baixa frequência. Possui três canais semicirculares e detecta vibrações a longas distâncias, podendo o tubarão se aperceber do som de um peixe a debater-se a uma distância de 250 a 600 m.
A linha lateral com poros e células eletrorreceptoras também é capaz de captar vibrações, correntes, alterações de temperatura e pressão da água. Sendo capaz até de localizar obstáculos e alimentos em água turvas. As células eletroreceptoras podem captar correntes elétricas produzidas por tecidos ativos de presas próximos.
Próximo aos poros há os neuromastos, células eletroreceptoras ciliadas que estão ligadas ao neurônio. Quando recebem um impulso elétrico, essas células se despolarizam levando a mensagem para o encéfalo.
Ainda como um sexto sentido, esses animais apresentam milhares de detectores especializados em seu focinho chamados ampolas de Lorenzini e são formados por uma rede de canais com electro-receptores cobertos uma substância gelatinosa. Quando há um estímulo externo a célula sensorial reage a partir de um pequeno potencial elétrico em sua membrana, levando os canais a permitir a entrada de íons de cálcio de carga positiva. O afluxo de carga positiva faz com que a célula libere neurotransmissores nas sinapses, ou pontos de contato, dos nervos para o cérebro, estimulando sua ativação. A taxa de estímulos indica a força e a polaridade do campo externo, enquanto sua localização relativa ao tubarão é supostamente determinada pela posição dos poros ativados em seu corpo. As células retornam ao seu estado original após a abertura de um segundo tipo de canal de membrana, que permite que a saída dos íons de potássio de carga positiva.
Para adentramos na sinalização celular, vamos relembrar um pouco sobre o sistema nervoso que é um tecido originado da placa neural, uma área diferenciada da ectoderme. Antes da 4º semana de gestação, a placa neural forma o tubo neural que sofre uma dilatação na região rostral ou anterior dando origem ao encéfalo. Nos pontos de encontro ou fechamento das extremidades da placa neural, no recém formado tubo neural, forma-se a crista neural que dá origem a componentes que a neuro-anatomia nomina como elementos periféricos e componentes celulares gliais.
O Sistema nervoso é dividido em sistema nervoso central (SNC), que está contido no canal vertebral e crânio, ou seja, o encéfalo e a medula espinhal e é onde as informações são recebidas e analisadas, ou seja, é o local onde ocorre as decisões e o envio de ordens; e sistema nervoso periférico (SNP), encontrado fora deste estojo ósseo, sendo os nervos (axônios) e gânglios e, carrega informações dos órgãos sensoriais para o sistema nervoso central e do sistema nervoso central para os órgãos efetores (músculos e glândulas).
Entretanto o sistema nervoso funcionalmente pode divide-se em somático, que lembra o sistema nervoso que atua em todas as relações que são percebidas por nossa consciência; e em visceral ou vegetativo aquele interage de forma inconsciente, no controle e na percepção do meio interno e vísceras. Tanto o somático quanto o egetativo, possuem componentes aferentes (sensitivos) e eferentes (motores).
No SNC, existem as chamadas substâncias cinzenta, formada pelos corpos dos neurônios, e substância branca, formada pelos prolongamentos dos neurônios. A unidade funcional e estrutural do sistema nervoso é o neurônio ou célula nervosa. São os neurônios que fazem a ligação entre as células receptoras dos diversos órgãos sensoriais e as células efetoras, nomeadamente músculos e glândulas. Os neurônios são células muito especializadas que apresentam um ou mais prolongamentos, ao longo dos quais se desloca um sinal elétrico e a função é receber, processar e enviar informações. Podem ser classificados, com base no sentido em que conduzem impulsos relativamente ao sistema nervoso central, em: neurônios sensoriais ou aferentes - os que transmitem impulsos do exterior para o sistema nervoso central; neurônios motores ou eferentes - os que transmitem impulsos do sistema nervoso central para o exterior; neurônios de conexão - os que conduzem impulsos entre os outros dois tipos de neurônios.
Os neurônios apresentam um corpo celular ou pericário que contém o núcleo e o citoplasma, onde estão contidos ribossomas, retículo endoplasmático granular e agranular e aparelho de Golgi, assim sintetiza todas as proteínas neuronais e realiza a maioria dos processos de degradação e renovação de constituintes celulares. Do corpo celular partem prolongamentos, denominado dendritos e axônios. Os dendritos apresentam as mesmas organelas citoplasmáticas do pericário. Traduzem os estímulos recebidos em alterações do potencial de repouso da membrana, que envolvem entrada e saída de determinados íons, causando pequenas despolarizações (excitatória) ou hiperpolarizações (inibitória). Os potenciais gerados nos dendritos se propagam em direção ao corpo e, neste, em direção ao cone de implantação do axônio.
O axônio é prolongamento longo e fino que pode ter origem de corpo ou de um dendrito principal, a partir de uma região denominada cone de implantação. Possui membrana plasmática, denominada axolema, e citoplasma, axoplasma. O axônio é capaz de gerar alteração de potencial de membrana (despolarização de grande amplitude) denominada potencial de ação ou impulso nervoso, e conduzi-lo até a terminação axônica, local onde ocorre a comunicação com outros axônios ou células efetuadoras. O local onde é gerado o impulso é chamado zona de gatilho. Esta especialização de membrana é devido à presença de canais de sódio e potássio, que ficam fechados no potencial de repouso, mas que se abrem quando despolarizações os atingem. Como os axônios não possuem ribossomas, toda a proteína necessária à manutenção destes deriva do pericário, e para que haja a renovação dos componentes das terminações é necessário um fluxo oposto, em direçãocão corpo. Esse fluxo de substâncias e organelas através do axoplasma é denominado fluxo axoplasmático.
Os neurônios são células altamente excitáveis que se comunicam entre si ou com células efetuadoras como por exemplo: células musculares, usando basicamente as alterações do potencial de membrana. A membrana celular separa o meio intracelular, onde predominam íons com cargas negativas e certa quantidade do íon potássio (K+), do meio extracelular, onde predominam cargas positivas, Sódio (Na+), Cálcio (Ca+) e certa quantidade do íon Cloro (Cl-). Essa diferença de cargas entre o meio intracelular e extracelular estabelece um potencial elétrico de membrana, que, como vimos no post anterior, é de aproximadamente -70mv, quando o neurônio está em repouso. Na membrana estão presentes canais iônicos seletivos, que se abrem ou fecham, permitindo a passagem de íons de acordo com o gradiente de concentração.
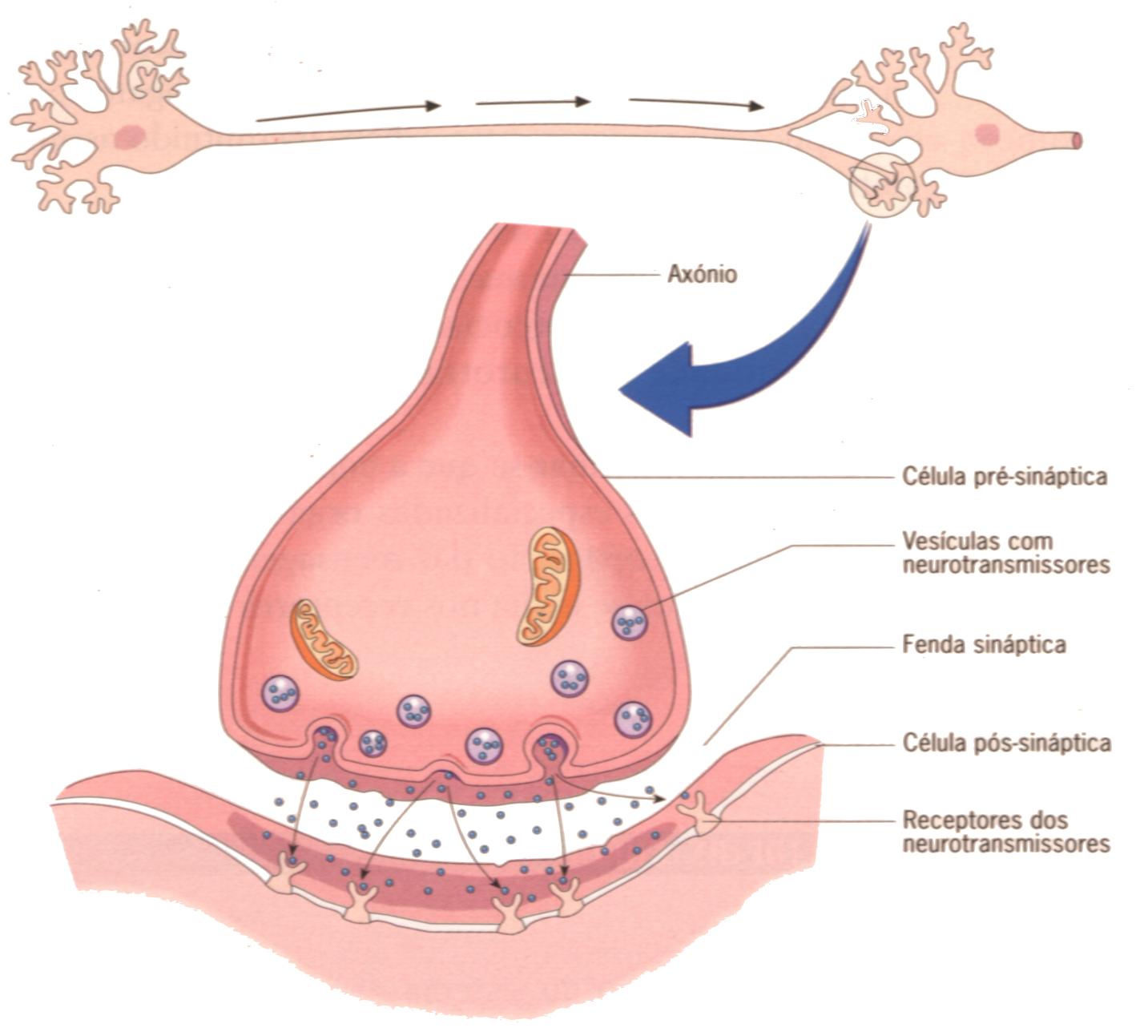
As fibras nervosas têm a função de propagar impulsos rápidos, e de transmiti-los à célula a próxima célula, através das sinapses. As sinapses podem acontecer entre dois neurônios, entre célula sensorial e neurônio ou entre neurônio e órgão efetor (músculo ou glândula). Assim, o impulso é captado pelos dendritos, passa ao corpo celular e deste para o axônio, que o envia para a célula seguinte.
No estado de repouso, o neurônio encontra-se polarizado, ou seja, o interior está carregado mais negativamente que o exterior. Ao atingir a membrana celular, o estímulo altera a permeabilidade aos íons Na+ e K+ no ponto excitado, permitindo assim, um influxo de íons sódio e a saída de íons potássio. Neste momento ocorre a despolarização, ou seja, diminui a negatividade no interior da célula. A entrada inicial de íons Na+ provoca a abertura de canais para esses íons nos segmentos seguintes, de modo que o processo se repete e o impulso nervoso se transmite através de todo o neurônio. Em alguns casos, a união de neurônios é tão estreita que a onda de despolarização passa diretamente do axônio de um neurônio a um dendrito do neurônio seguinte, o que se denomina sinapse elétrica. As sinapses elétricas são exclusivamente entre neurônios e raras em vertebrados. A comunicação entre dois neurônios se dá através de canais iônicos presentes em cada uma das membranas em contato, que permitem a passagem direta de pequenas moléculas do citoplasma de uma das células para o da outra.
Nas sinapses químicas, que ocorre mais frequentemente, o sinal elétrico que chega à terminação axônica, provoca a liberação de neurotransmissores (mensageiros químicos presentes no interior de vesículas na terminação axônica). Ao atingir a terminação axônica, o potencial de ação faz com que as vesículas se fusionem com a membrana da terminação, liberando os neurotransmissores que estavam contidos para a fenda sináptica (espaço virtual entre o neurônio e a célula efetora). Ao serem liberados na fenda sinóptica, os neurotransmissores se ligam a receptores específicos presentes na membrana da célula pós-sináptica (célula efetora). A ligação do neurotransmissor com o seu receptor específico, gera uma alteração no potencial de membrana da célula efetora, transmitindo o impulso nervoso e gerando uma resposta . Podemos então concluir que a transmissão do impulso implica a transformação de um sinal elétrico em um sinal químico que, posteriormente, é transformado em um outro sinal elétrico. A sinapse química sempre apresenta um elemento pré-sináptico, que armazena e libera o neurotransmissor, um elemento pós-sináptico, que contém o receptor, e uma fenda sináptica, que separa as duas membranas. Desta maneira, na célula pré-sinaptica, há a membrana pré-sináptica, que apresenta projeções densas que mantém de forma organizada as vesículas sinápticas e na célula pós-sináptica, há a membrana pós-sináptica, que possui os receptores específicos para os neurotransmissores.
Os axônios são cobertos por uma membrana denominada bainha de mielina, que possui a característica de isolante elétrico, impedindo que as cargas elétricas se dispersem. Assim, condução do impulso nervoso nas fibras mielínicas (com bainha de mielina) e amielínicas (sem bainha de mielina) difere na sua velocidade, sendo maior nas mielínicas. No trajeto do axônio, há regiões chamadas nódulos de Ranvier, em que a bainha de mielina é interrompida, gerando assim a condução saltatória, nos quais o impulso nervoso é transmitido, aos saltos, de um nódulo de Ranvier ao outro, ao longo da fibra (axônio).
Quando um impulso nervoso atinge a membrana pré-sináptica há uma alteração no potencial, estimulando a liberação do neurotransmissor na fenda sináptica, que atinge os receptores da célula pós-sináptica. Estes receptores podem ser canais iônicos que se abrem quando em contato com o neurotransmissor, permitindo a entrada ou saída de determinados íons. A movimentação de íons, tanto para dentro, quanto para fora, causa alterações no potencial de membrana. Quando o receptor não é um canal iônico, a sua combinação com o neurotransmissor gera uma nova molécula chamada de segundo mensageiro, que causará modificações na célula pós-sináptica. Após o contato com o receptor, é necessário que o neurotransmissor seja removido da fenda sináptica para que não haja excitação ou inibição por tempo prolongado. Essa remoção pode ser feita por ação enzimática ou por recaptação pela membrana pré-sináptica; e uma vez dentro da terminação nervosa, o neurotransmissor pode ser reutilizado ou inativado.
Endorfinas e encefalinas: bloqueiam a dor, agindo naturalmente no corpo como analgésicos.
Dopamina: neurotransmissor inibitório derivado da tirosina. Produz sensações de satisfação e prazer.
Serotonina: neurotransmissor derivado do triptofano regula o humor, o sono, a atividade sexual, o apetite, as funções neuroendócrinas, temperatura corporal, sensibilidade à dor, atividade motora e funções cognitivas.
GABA (ácido gama-aminobutirico): principal neurotransmissor inibitório do SNC. Ele está presente em quase todas as regiões do cérebro, embora sua concentração varie conforme a região. Está envolvido com osprocessos de ansiedade. Seu efeito ansiolítico seria fruto de alterações provocadas em diversas estruturasdo sistema límbico, inclusive a amígdala e o hipocampo. A inibição da síntese do GABA ou o bloqueio de seus neurotransmissores no SNC, resultam em estimulação intensa, manifestada através de convulsõesgeneralizadas.
Ácido glutâmico ou glutamato: principal neurotransmissor estimulador do SNC. A sua ativação aumenta a sensibilidade aos estímulos dos outros neurotransmissores.


















.jpg)






{kind=link}
{kind=link}
{kind=link}